神経終末端におけるシナプス小胞のエンドサイトーシスはシナプス伝達の維持に不可欠であるが、その分子メカニズムには不明な点が多く残されている。そこで我々は、哺乳類聴覚系の中継核である台形体内側核に存在する巨大神経終末端の膜容量を測定することにより、シナプス小胞のエンドサイトーシスの分子メカニズムを調べた。ボツリヌス毒素Eを神経終末端内に投与してシナプス伝達を停止させると、従来“kiss-and-run”型の開口放出を反映するとみなされていた時定数1秒以下の速い膜容量変化だけが残存し、これは神経伝達物質の放出に関わらないと判定された。伝達物質開口放出に関わる膜容量変化のみを解析すると、小胞エンドサイトーシスに伴う膜容量変化速度の時定数は10~25秒であり、放出量が増大するにつれ遅くなることが明らかになった。また、加水分解を受けないGTPアナログGTPgS、またはGTP分解酵素ダイナミン1のプロリン・リッチ・ドメイン(PRD)に相当するペプチドを神経終末端内に投与してダイナミン1の作用を阻止すると、エンドサイトーシスが停止した。これらは開口放出に対する即時的な効果を持たなかったが、開口放出を繰り返すと開口回数に依存して放出量を減少させる効果を示した。したがって、神経終末端において、ダイナミン1によるGTPの加水分解が小胞のエンドサイトーシスとそれによるシナプス伝達の維持に必要不可欠であると結論された。(Science, 307: 124-127, 2005)
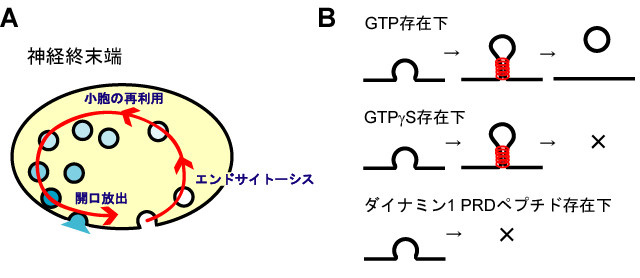
図の説明
A: 神経終末端では開口放出によって使われたシナプス小胞膜は、エンドサイトーシスによって回収され、再利用される。
B: シナプス小胞のエンドサイトーシスにはダイナミン1(赤)によるGTP加水分解が必須である。GTPgS存在下ではダイナミン1によるGTP加水分解に伴う小胞形成が阻害され、ダイナミン1 PRDペプチド存在下ではエンドサイトーシス部位へのダイナミン1の動員が阻害されると考えられる。