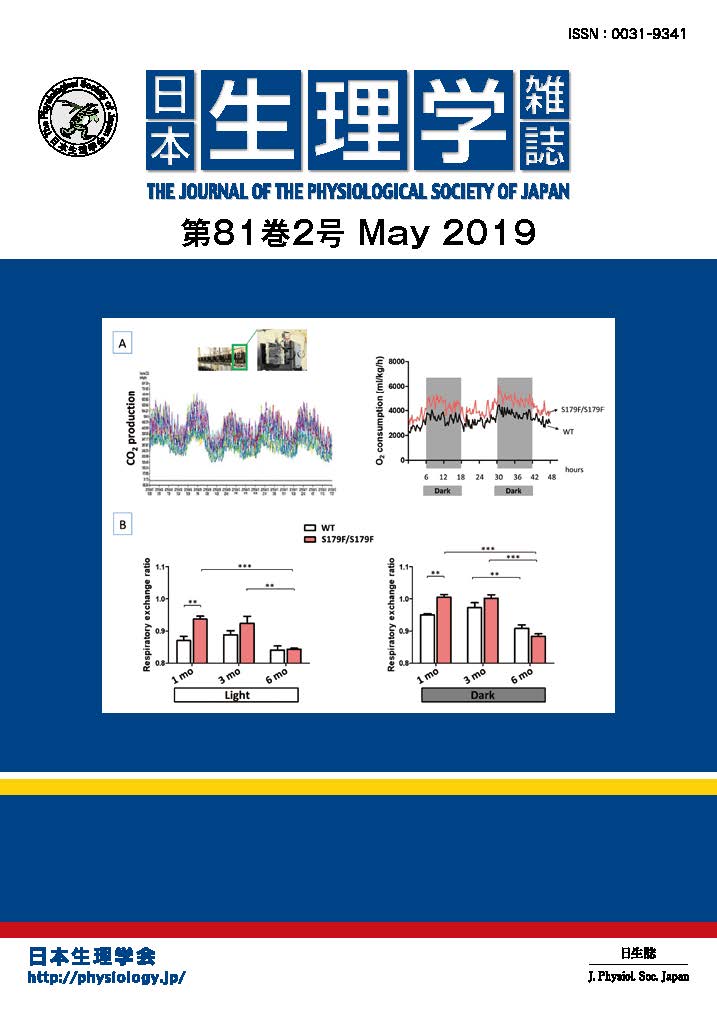
海馬は「いつ、どこで、何があったか」というエピソード記憶の形成に中心的な役割を持ちます。海馬には時間や空間の情報が入り、特定のエピソードに反応するニューロンも発見されました。しかし、海馬内に記憶情報がどの様に記録されるか、その様態はよく解っていません。
ラットを用いて、特定のエピソードを学習させる回避学習実験を行うと、AMPA受容体がCA1シナプスに移行して学習が成立します(PNAS 2011)。しかし、何がAMPA受容体をシナプスに移行させ、興奮性シナプスを強化するか、その引き金分子は不明でした。本研究ではまずアセチルコリンに着目し、学習前後の海馬CA1領域内の分泌量を測定しました。すると、アセチルコリンは学習中から分泌量が増加し、学習後にも高く維持されることが明らかとなりました。さらに、スライスパッチクランプ法を用い、個々の海馬CA1ニューロンについてシナプス機能を解析しました。すると回避学習は、AMPA受容体を介する興奮性シナプスの可塑性を高めるだけでなく、GABAA受容体を介した抑制性シナプスの可塑性を高める結果、個々のCA1ニューロンが複雑かつ多様なシナプス入力を保持することが明らかになりました(図)。興奮と抑制、どちらのシナプス変化も学習成立に必要であり、アセチルコリン受容体の一種である「ムスカリン性 M1受容体」を阻害すると興奮性シナプスの可塑性が阻害され、回避学習ができなくなりました。一方、「ニコチン性 α7受容体」を阻害すると抑制性シナプスの可塑性が阻害され、回避学習ができなくなりました。本研究から、興奮性シナプスと抑制性シナプスの多様な可塑性により記憶情報が符号化され、エピソード記憶が成立すると考えられます。
(Mitsushima et al, Nature Communications 4:2760 doi: 10.1038/ ncomms3760, 2013)
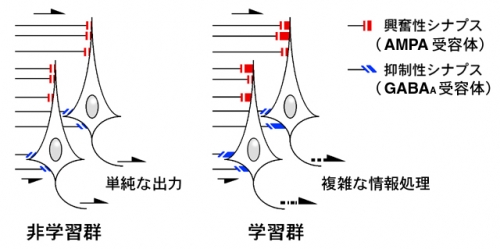
図 非学習群と学習群における、海馬CA1ニューロンとシナプスの変化。矢印は情報の流れを示す。
学習群ではアセチルコリンが引き金となり、AMPA受容体を介する興奮性シナプスだけでなく、GABAA受容体を介する抑制性シナプスも強化されることが判明した。結果、個々のニューロンに特徴的な出力が形成され、複雑な記憶の情報処理が可能になると考えられる。