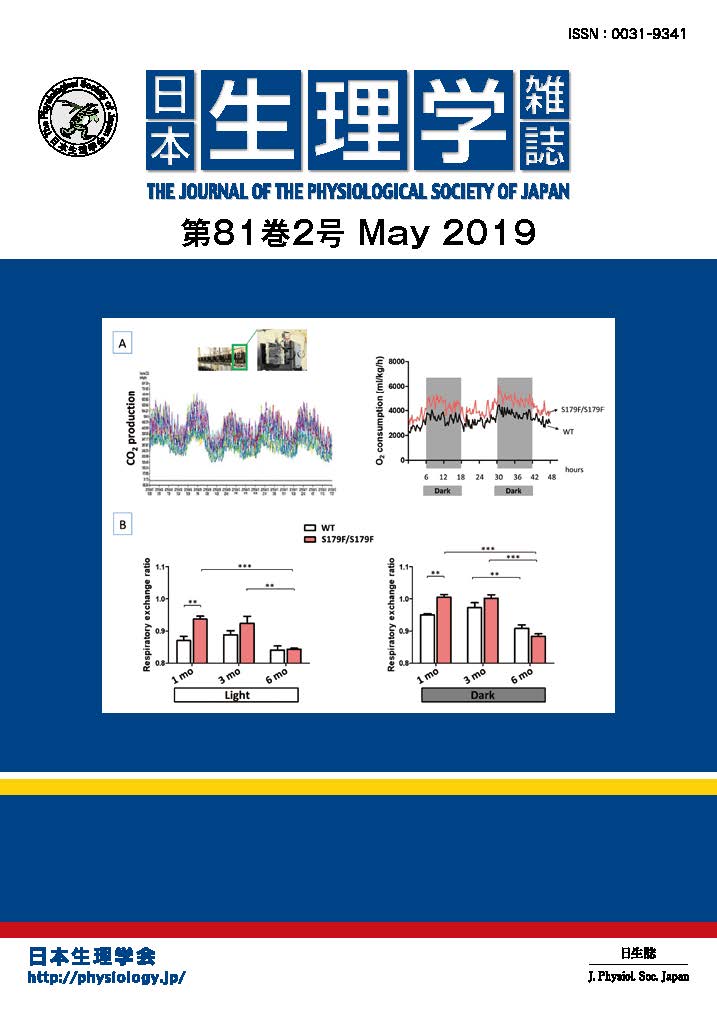
心筋細胞の興奮と収縮には, 細胞膜に存在するL型Ca2+チャネルを通して細胞内へカルシウムイオン(Ca2+)が流入することが重要です. 交感神経終末部から遊離される神経伝達物質ノルアドレナリンは, 心筋の細胞膜に存在するα1及びβアドレナリン受容体を刺激して, Ca2+チャネルの機能を調節しています. β受容体を介したCa2+チャネルの制御機構は, 以前より詳細に研究がなされてきましたが, α1受容体刺激によってCa2+チャネルが, どのようなメカニズムで調節されているのか, 近年まで明らかとなっていませんでした. 今回, 私たちは, ラット心室筋細胞を用いて, α1受容体を刺激したときにCa2+チャネルがどのようなメカニズムで調節されているのか, 詳細を明らかにしました. 本研究により,心筋細胞膜にはα1A及びα1B受容体という異なった二種類のα1受容体があり, それぞれの受容体を刺激すると, 複雑な情報伝達経路を介してCa2+チャネルに働きかけ, Ca2+流入量を増加、あるいは減少するという正反対のはたらきを示すことが初めてわかりました. 今回, 私たちは, α1アドレナリン受容体刺激が心臓の機能を調節しているメカニズムの一部を解明しましたが, これが基礎となり, α1アドレナリン受容体刺激による心臓生理機能の制御メカニズムの全容が解明されるものと考えています(O-Uchi J et al., Proc Natl Acad Sci U S A. 102:9400-9405, 2005. O-Uchi J et al., Circ Res. 102: 1378-1388, 2008).
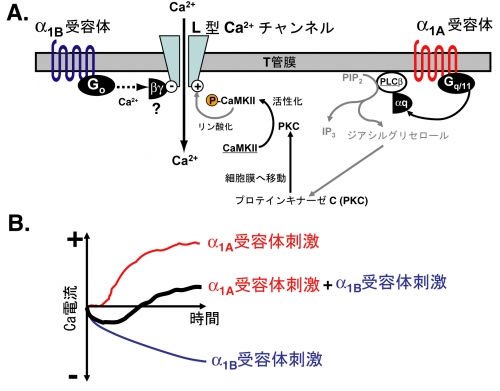
図の説明
心筋α1アドレナリン受容体シグナルによるL型Ca2+チャネルの制御メカニズム
心室筋細胞には、α1A及びα1Bアドレナリン受容体という二種類のα1受容体が存在し, それぞれの受容体を刺激すると, 異なる細胞情報伝達系を介してCa2+チャネルに対して正反対のはたらきを及ぼす(図A)。α1A受容体を刺激するとGqタンパク質を介し, プロテインキナーゼC (PKC)やCa2+/カルモジュリン依存性キナーゼII (CaMKII)などの酵素が活性化され, Ca2+流入量(Ca2+電流)が増加する(図B, 赤線). 一方, α1B受容体は別の種類のGタンパク質(Go)と結合していて, この経路がCa2+チャンネルに抑制的に作用し, Ca2+流入量を減少させることがわかった (図B, 青線). 二つの受容体が同時に刺激されると、二つの細胞情報伝達系の働きの合算として、Ca2+流入量の時間経過は、特徴的な二相性変化を示す(図B, 黒線).