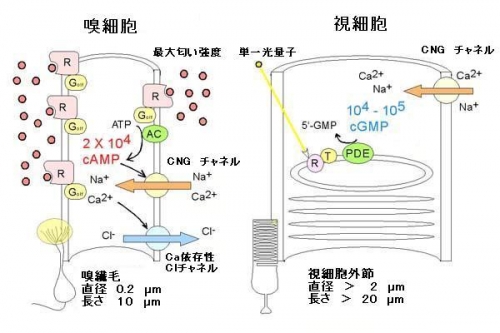
本研究では、「香り」を受け取る際の、私達の体の中での信号増幅の機構を明らかにしました。香りを受け取る場所は直径200nmと いう微細な構造体(嗅繊毛)であるために、これまでこの課題を解くのは実験的に困難であるとされてきました。今回、我々は新しい研究手法として光解離物質 と電気生理学的手法を合わせることで、実験的な困難を克服し、微細な神経プロセス内で二次伝達物質(セカンドメッセンジャー)として働くcAMPの実時間 濃度測定を可能としました。感覚細胞としては、既に同様なG蛋白介在性のシステムを用いる視細胞(大きな感覚細胞)での信号増幅の様式が詳しく調べられて います。視細胞ではたった1つの光の粒(光量子=フォトン)が酵素過程による信号増幅により1万-10万分子のセカンドメッセンジャー(cGMP)を加水 分解し、これによって視細胞が一光量子に応答することが一般的に知られています。しかし、今回の新しい技術で調べてみると、嗅細胞では最大強度の匂いが到 達した場合でも、cAMPの生成は繊毛1本全体で2万分子/秒程度であり、視細胞での増幅よりも非常に小さいことが分かりました。一見、このシステムは非 効率に見えますが、低増幅の酵素システムはATP(エネルギー源)の消費を最低限に抑えるという点で非常に合理的であると言えます。では、嗅細胞では、如 何にして信号増幅をしているのでしょうか?実は、繊毛という微細構造体は、大きな表面積/容量比を持つことによって、わずかなセカンドメッセンジャー分子 (cAMP)の変化を増幅しています。小さな容量内では、わずかな分子変化が大きな濃度変化につながります。更に、嗅細胞では、視細胞にはない「サード メッセンジャー」とも呼ぶべきCaイオンが興奮性のClチャネルを開口することで更なる信号増幅を行うことも明らかになりました。このように、生体の細胞 は目的に合わせて、分子の役割を少しずつ調節しているのでしょう(Journal of Neuroscience, 2005
Nov 30;25(48):11084-91.)。
嗅 細胞では最大の匂い刺激を行った場合、1繊毛当たり2万分子のcAMPが生成するのみであるが、視細胞では1つのphotonで刺激した場合、1-10万 分子のcGMPが分解する。両者の分子カスケードの構成は非常によく似ているが、細胞の形態(表面積/容量比)、生物学的戦略(Caイオンによる付加的増 幅回路)によって、そのシステムは大きく異なっている。略語は以下の通り、R:リセプタ蛋白、Golf:嗅覚特異性G蛋白質、AC:アデニレートシクラー ゼ、T:トランスデューシン、PDE:ホスホジエステラーゼ。